Feature Review
Evolutionary Dynamics of Maize: Implications from Gene Flow Studies 
 Author
Author  Correspondence author
Correspondence author
Maize Genomics and Genetics, 2024, Vol. 15, No. 4 doi: 10.5376/mgg.2024.15.0018
Received: 07 Jun., 2024 Accepted: 15 Jul., 2024 Published: 30 Jul., 2024
Wang D.L., and Huang Y.C., 2024, Evolutionary dynamics of maize: implications from gene flow studies, Maize Genomics and Genetics, 15(4): 182-190 (doi: 10.5376/mgg.2024.15.0018)
This study explores the evolutionary dynamics of maize, focusing on the impact of gene flow on its genetic diversity and adaptation. The study analyzes various aspects of gene flow, including its role in altering genetic diversity, influencing agronomic traits, and affecting pest and disease resistance. Key findings indicate that gene flow from modern maize varieties to traditional landraces and wild relatives significantly contributes to genetic diversity and the development of traits that enhance adaptability and resilience. The integration of beneficial alleles through gene flow facilitates maize's adaptation to diverse ecological niches and changing environmental conditions. Additionally, this study discusses the implications of gene flow for maize breeding and conservation strategies, highlighting the need for careful management to harness its benefits while mitigating potential adverse effects on indigenous maize varieties and their ecosystems. Future directions emphasize the importance of integrating ecological and genomic data to inform sustainable breeding programs and conservation efforts, ensuring maize remains a vital global staple in response to global challenges like climate change.
1 Introduction
Maize (Zea mays L.) is one of the most significant cereal crops globally, serving as a primary source of food, feed, and industrial products. Its importance is underscored by its extensive cultivation and high yield, particularly in regions like the US central Corn Belt, where yields have increased dramatically over the past century due to advancements in breeding and agricultural practices (Kong et al., 2020). Originating in Mexico and Central America, maize has a rich history of domestication and cultivation, making it a staple crop in many parts of the world (Lal et al., 2020).
The evolutionary dynamics of crops involve complex interactions between genetic variation, environmental factors, and human-mediated selection. In maize, these dynamics are evident in the remarkable phenotypic diversity and adaptability of different genotypes. Studies have shown that maize's genetic makeup includes over 32 000 genes, which contribute to its adaptability and resilience (Lal et al., 2020). The evolutionary history of maize is marked by multiple waves of domestication and gene flow, which have shaped its current genetic landscape (Kistler et al., 2020).
Gene flow studies are crucial for understanding the evolutionary history and genetic diversity of maize. These studies reveal how genetic material has been exchanged between different maize populations over time, contributing to the development of new varieties and enhancing genetic diversity. For instance, archaeological evidence suggests that ancient gene flow from South America to Central America played a significant role in the development of more productive maize varieties, which in turn supported the growth of complex societies in these regions (Kistler et al., 2020). Additionally, comparative genomic studies highlight the intraspecies variation in maize, further emphasizing the importance of gene flow in shaping its genetic structure (Haberer et al., 2020).
This study aims to explore the evolutionary dynamics of maize through the lens of gene flow studies. By examining the historical and contemporary patterns of gene flow, this study aims to provide insights into how genetic exchanges have influenced maize's development and adaptation. Understanding these dynamics can inform future breeding programs and contribute to the sustainable improvement of maize as a vital agricultural resource.
2 Background on Maize Evolution
2.1 Origin and domestication of maize
Maize (Zea mays ssp. mays) is one of the most important crops globally, with its domestication tracing back approximately 9 000 years to the Balsas Basin in southwestern Mexico. The domestication process involved the transformation of the wild grass teosinte (Zea mays ssp. parviglumis) into the cultivated maize we know today. Recent genomic studies have suggested that the domestication of maize may have involved multiple events and significant gene flow from other teosinte subspecies, such as Zea mays ssp. mexicana, challenging the earlier consensus of a single domestication event (Kistler et al., 2020; Moreno-Letelier et al., 2020). Additionally, archaeological evidence indicates that maize was dispersed to South America as a partial domesticate, where it underwent further domestication and improvement before being reintroduced to Central America (Kistler et al., 2020).
2.2 Genetic diversity in wild and cultivated maize
The genetic diversity of maize is vast, encompassing a wide range of landraces and modern varieties. This diversity results from both ancient and contemporary gene flow events between wild relatives and cultivated maize. Studies have shown that the maize genome contains over 103 000 pan-genes, with significant variation in gene content, genome structure, and DNA methylation patterns across different maize lines (Hufford et al., 2020; Xu et al., 2020). Figure 1A and 1C illustrate these variations in the gene models categorized by different phylostrata and the distribution of core, near-core, dispensable, and private genes, demonstrating the genetic diversity and structural complexity of the maize genome. The genetic architecture of maize has been shaped by both natural selection and human-mediated breeding, leading to the development of traits beneficial for various environments and agricultural practices (Chen et al., 2020; Hu et al., 2022). The introgression of genes from teosinte has played a crucial role in maize's adaptation to diverse ecological niches, including highland regions (Calfee et al., 2021; Hu et al., 2022), as supported by the expression profiles in different tissues shown in Figure 1D of the diagram (Figure 1). This substantiates the significant genetic diversity present in maize, which is crucial for its adaptation and resilience across various environments.
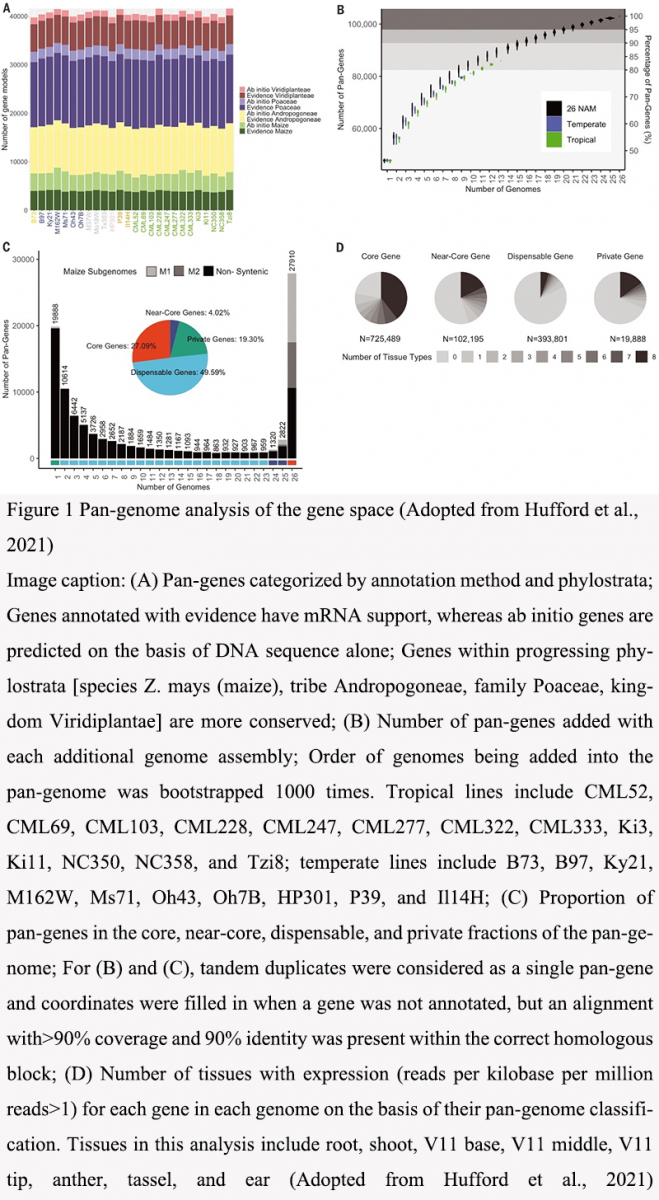 Figure 1 Pan-genome analysis of the gene space (Adopted from Hufford et al., 2021) Image caption: (A) Pan-genes categorized by annotation method and phylostrata; Genes annotated with evidence have mRNA support, whereas ab initio genes are predicted on the basis of DNA sequence alone; Genes within progressing phylostrata [species Z. mays (maize), tribe Andropogoneae, family Poaceae, kingdom Viridiplantae] are more conserved; (B) Number of pan-genes added with each additional genome assembly. Order of genomes being added into the pan-genome was bootstrapped 1000 times; Tropical lines include CML52, CML69, CML103, CML228, CML247, CML277, CML322, CML333, Ki3, Ki11, NC350, NC358, and Tzi8; temperate lines include B73, B97, Ky21, M162W, Ms71, Oh43, Oh7B, HP301, P39, and Il14H. (C) Proportion of pan-genes in the core, near-core, dispensable, and private fractions of the pan-genome. For (B) and (C), tandem duplicates were considered as a single pan-gene and coordinates were filled in when a gene was not annotated, but an alignment with>90% coverage and 90% identity was present within the correct homologous block; (D) Number of tissues with expression (reads per kilobase per million reads>1) for each gene in each genome on the basis of their pan-genome classification. Tissues in this analysis include root, shoot, V11 base, V11 middle, V11 tip, anther, tassel, and ear (Adopted from Hufford et al., 2021) |
.png){kind=link}
2.3 Historical perspectives on gene flow in maize
Gene flow has been a significant factor in the evolutionary history of maize. Hybridization between maize and its wild relatives, such as teosinte, has introduced genetic variation that has facilitated maize's adaptation to new environments and contributed to its global spread. For instance, introgression from the highland teosinte Zea mays ssp. mexicana has been crucial for maize's adaptation to high-altitude regions in Mexico (Calfee et al., 2021). Additionally, the backflow of genetic material from South American maize varieties to Central America has been hypothesized to enhance the genetic diversity and productivity of maize in these regions (Kistler et al., 2020). The selective sorting of ancestral introgression along environmental gradients further highlights the complex interplay between gene flow and local adaptation in maize evolution (Calfee et al., 2021).
3 Methodologies in Gene Flow Studies
3.1 Genetic markers and molecular tools
Genetic markers and molecular tools are fundamental in studying gene flow in maize. Techniques such as next-generation sequencing (NGS) have revolutionized the field by enabling rapid and comprehensive sequencing of plant genomes. For instance, the use of Illumina sequencing to analyze nearly complete genomes of Maize dwarf mosaic virus (MDMV) isolates has provided insights into genetic diversity and evolutionary relationships (Wijayasekara et al., 2021). Additionally, genotyping by target sequencing (GBTS) technology has been employed to develop low-density genotyping platforms, such as the 5.5 K SNP markers panel, which are crucial for genetic and molecular breeding studies (Ma et al., 2022). These tools facilitate the identification of genetic variations and the mapping of quantitative traits, thereby enhancing the understanding of gene flow dynamics in maize.
3.2 Population genomics
Population genomics involves the study of genetic variations within and between populations of maize. High-quality genomic sequences from diverse maize lines have been produced to map important traits and demonstrate the genetic diversity of maize (Hufford et al., 2021). This approach has revealed significant variation in gene content, genome structure, and DNA methylation across different maize accessions. By leveraging whole-genome sequencing (WGS) and whole genome bisulfite sequencing (WGBS), researchers have investigated the adaptive and phenotypic consequences of methylation variations in maize populations, providing insights into the evolutionary forces acting on DNA methylation patterns (Xu et al., 2020). These population genomics studies are essential for understanding the extent and impact of gene flow in maize.
3.3 Phylogeographic approaches
Phylogeographic approaches combine phylogenetic and geographic data to study the historical processes that have shaped the genetic structure of maize populations. Phylogenetic analysis based on complete genome sequences has been used to reveal evolutionary relationships among MDMV isolates, highlighting the divergence of isolates from different regions (Wijayasekara et al., 2021). Such analyses help trace the movement and spread of genetic variants across geographical landscapes, providing a historical context for gene flow events. By integrating phylogenetic data with geographic information, researchers can infer the origins and migration patterns of maize populations, thereby elucidating the evolutionary dynamics of gene flow.
3.4 Experimental and observational studies
Experimental and observational studies are crucial for validating hypotheses about gene flow in maize. For example, the development and validation of the 5.5 K SNP markers panel using GBTS technology involved extensive genetic analyses of two maize populations, revealing genetic divergences and prediction accuracies for various traits (Ma et al., 2022). These studies provide empirical data on the genetic structure and diversity of maize populations, which are essential for understanding gene flow. Observational studies, such as the analysis of DNA methylation patterns in modern maize, landrace, and teosinte populations, have also shed light on the role of epigenetic variations in adaptive evolution (Xu et al., 2020). Together, these experimental and observational approaches contribute to a comprehensive understanding of gene flow dynamics in maize.
4 Patterns of Gene Flow in Maize
4.1 Gene flow between wild relatives and cultivated maize
Gene flow between wild relatives and cultivated maize is a critical factor in the evolutionary dynamics of maize. Wild relatives of crops, such as teosinte, serve as reservoirs of genetic diversity that can be introgressed into cultivated maize. This gene flow can introduce beneficial traits, such as disease resistance and environmental adaptability, into the cultivated gene pool. For instance, the study on crop wild relatives (CWRs) emphasizes the importance of capturing genetic variation from wild relatives to enhance crop improvement efforts (Egan et al., 2018). Similarly, research on wheat has shown that introgression from wild relatives significantly contributes to the adaptive diversity of modern crops, suggesting a parallel importance in maize.
4.2 Introgression and hybridization events
Introgression and hybridization events play a pivotal role in shaping the genetic landscape of maize. These processes involve the incorporation of genetic material from one species into the gene pool of another through repeated backcrossing. The study on wheat highlights how introgression from wild relatives has historically contributed to the adaptive evolution of crops by increasing genetic diversity and reducing deleterious alleles (He et al., 2019). This phenomenon is likely mirrored in maize, where hybridization with wild relatives can introduce new alleles that enhance agronomic traits and environmental adaptability.
4.3 Spatial and temporal patterns
The spatial and temporal patterns of gene flow in maize are influenced by various factors, including geographic isolation and environmental conditions. The research on woodland strawberry demonstrates that landscape isolation and mesoclimatic variation are significant determinants of genetic variation in wild populations (Egan et al., 2018). These findings suggest that similar spatial proxies could be used to predict gene flow patterns in maize. Temporal patterns of gene flow are also crucial, as they can indicate historical introgression events and ongoing hybridization processes that shape the genetic structure of maize populations over time.
4.4 Impact of human activities on gene flow
Human activities have a profound impact on gene flow in maize. Agricultural practices, such as the cultivation of genetically modified (GM) crops and the movement of seeds across regions, can facilitate or hinder gene flow between wild and cultivated populations. The study on CWRs advocates for the use of evolutionary approaches to capture and conserve genetic variation, highlighting the role of human intervention in managing gene flow (Egan et al., 2018). Additionally, the research on wheat underscores the importance of human-mediated selection in shaping the adaptive landscape of crops through introgression (He et al., 2022). These insights underscore the need for careful management of gene flow to balance crop improvement with the conservation of genetic diversity.
5 Implications of Gene Flow
5.1 Genetic diversity and adaptation
Gene flow plays a crucial role in maintaining and enhancing genetic diversity within maize populations. The introduction of alleles from modern varieties (MVs) into traditional landraces (LRs) and wild relatives (WRs) has been shown to increase genetic diversity, which is essential for adaptation to changing environmental conditions. For instance, a study demonstrated that gene flow from MVs into LRs and WRs in Mexico led to an increase in genetic diversity post-2000, likely due to the introgression of beneficial alleles from MVs (Rojas-Barrera et al., 2019). This enhanced genetic diversity is vital for the evolutionary capacity of maize populations to adapt to novel environments, as evidenced by the retention of 99% of the standing molecular marker variation in a population subjected to directional selection for flowering time adaptation (Wisser et al., 2019).
5.2 Evolution of agronomic traits
Gene flow significantly influences the evolution of agronomic traits in maize. The introgression of alleles from MVs into LRs and WRs can lead to rapid changes in traits such as flowering time, yield, and drought resistance. For example, a high-resolution genome-wide association study identified hundreds of candidate genes associated with genotype by environment interactions, which are crucial for agronomic performance across diverse environments (Gates et al., 2019). Additionally, the domestication process of maize, which involved the transformation of teosinte into modern maize, was driven by genetic interactions and selection pressures that shaped key agronomic traits (Stitzer et al., 2018). These findings underscore the importance of gene flow in the ongoing evolution and improvement of agronomic traits in maize.
5.3 Pest and disease resistance
Gene flow also has implications for pest and disease resistance in maize. The introduction of alleles from MVs into LRs and WRs can enhance resistance to pests and diseases, which is critical for crop sustainability. The genomic evidence of ongoing evolution in maize due to introgression from MVs suggests that beneficial alleles conferring resistance to biotic stresses may be incorporated into the genetic pool of LRs and WRs (Rojas-Barrera et al., 2019). This process can lead to the development of maize varieties with improved resistance to pests and diseases, thereby reducing the reliance on chemical control methods and promoting sustainable agricultural practices.
5.4 Conservation and biodiversity
The conservation of genetic diversity and biodiversity in maize is closely linked to gene flow. The introgression of alleles from MVs into LRs and WRs not only enhances genetic diversity but also contributes to the conservation of these genetic resources. Monitoring strategies and policies that promote the use and safeguarding of genetic diversity are essential for the conservation of maize at its center of origin (Rojas-Barrera et al., 2019). Furthermore, understanding the evolutionary dynamics of maize populations, including the role of gene flow, is crucial for developing effective conservation strategies that ensure the long-term sustainability of maize biodiversity (Stitzer et al., 2018; Rojas-Barrera et al., 2019; Gates et al., 2019).
6 Case Studies
6.1 Gene flow in modern agricultural landscapes
Gene flow in modern agricultural landscapes has been a significant factor influencing the genetic diversity and evolution of maize. Contemporary studies have shown that the introduction of modern maize varieties (MVs) into traditional agricultural systems can lead to rapid genetic changes in indigenous landraces (LRs) and crop wild relatives (WRs). For instance, research conducted in Mexico, the center of origin for maize, demonstrated that gene flow from MVs has resulted in notable genomic changes in both LRs and WRs over the past 60 years. This gene flow has led to increased genetic diversity in LRs and a decrease in genetic divergence between MVs and both LRs and WRs, highlighting the dynamic nature of maize evolution in response to agricultural practices (Rojas-Barrera et al., 2019). Figure 2 illustrates the clustering of modern varieties (MV1 and MV2) with landraces collected during different periods (earlier than 1960, between 1960 and 1980, and later than 2000) and their genetic interactions as revealed by genotyping data. The clustering patterns shown in panels Figure 2A and 2B of the figure reflect the genetic integration of these maize varieties across different time frames and geographical distributions, which supports the study’s findings on the impact of gene flow on maize diversity and its adaptability to various agricultural environments (Rojas-Barrera et al., 2019).
 Figure 2 MVs clustering with germplasm from different breeding programs (Adopted from Rojas-Barrera et al., 2019) Image caption: (A): Genotype distribution of MV1 and MV2 sampled in sympatry with LRs. LRs are colored by sampling period: earlier than 1960 (LR<1960), dark blue; between 1960 and 1980 (LR 1960 to 1980); light blue; and later than 2000 (LR>2000), purple; (B): Distribution for MV1, MV2, and the LRs collected for this work with a sample subset from the US national maize inbred seed bank (2 578 genotypes and 13 953 SNPs); (C): Clustering of genotypes colored by the breeding program (1 002 genotypes, 13 953 SNPs); ExPVP, expired plant variety protection. (D) Tropical breeding pools from Mexico, Nigeria, Cameroon, MV1, and MV2 (463 genotypes and 13 953 SNPs) (Adopted from Rojas-Barrera et al., 2019) |
.png){kind=link}
6.2 Impact of transgenic maize on wild relatives
The impact of transgenic maize on wild relatives is a critical area of study, particularly concerning the potential for gene flow and its ecological consequences. Transgenic maize varieties, designed for traits such as pest resistance and herbicide tolerance, can cross-pollinate with wild relatives, leading to the introgression of transgenes into wild populations. This gene flow can alter the genetic makeup of wild relatives, potentially affecting their fitness and ecological roles. Studies have shown that gene flow from transgenic maize to wild relatives like teosinte can occur, necessitating careful monitoring and management strategies to preserve the genetic integrity and ecological functions of wild maize relatives (Rojas-Barrera et al., 2019).
6.3 Adaptive introgression in different environments
Adaptive introgression, the incorporation of beneficial alleles from one population into another through hybridization, plays a crucial role in the evolutionary dynamics of maize. Research has demonstrated that maize populations can rapidly adapt to new environments through the introgression of advantageous alleles. For example, a study on a tropical landrace of maize translocated to a temperate environment revealed significant genomic shifts over ten generations of selection for earlier flowering time. This adaptive introgression allowed the population to achieve a 26-day reduction in flowering time while maintaining high levels of genetic diversity, showcasing the potential for rapid environmental adaptation in maize (Wisser et al., 2019).
6.4 Comparative studies in other crops
Comparative studies in other crops provide valuable insights into the evolutionary dynamics of gene flow and adaptation. For instance, research on crop wild relatives (CWRs) of woodland strawberry (Fragaria vesca) has highlighted the importance of natural selection, gene flow, and genetic drift in shaping genetic variation. These studies emphasize the need for an evolutionary approach to capture and conserve genetic diversity in CWRs, which can be applied to other crops like maize. By understanding the evolutionary processes in different crops, researchers can develop strategies to enhance crop resilience and adaptation to changing environments (Egan et al., 2018).
7 Future Directions
7.1 Advances in genomic technologies for gene flow studies
The rapid evolution of genomic technologies has significantly enhanced our ability to study gene flow in maize. High-throughput sequencing and advanced genotyping methods, such as genotyping by sequencing (GBS), have provided detailed insights into the genetic changes occurring in maize populations over time. For instance, the use of GBS has revealed ongoing introgression from modern maize varieties (MVs) into landraces (LRs) and wild relatives (WRs), highlighting the dynamic nature of maize genomes in response to gene flow (Rojas-Barrera et al., 2019). Future advancements in genomic technologies, including more refined sequencing techniques and bioinformatics tools, will likely provide even deeper insights into the mechanisms and consequences of gene flow in maize.
7.2 Integrative approaches combining ecology and genomics
Combining ecological data with genomic information offers a comprehensive understanding of how gene flow impacts maize evolution. Studies have shown that the geographical distribution and environmental adaptation of maize are influenced by both genetic and ecological factors (Wisser et al., 2019). Integrative approaches that merge ecological modeling with genomic data can elucidate how environmental pressures shape genetic diversity and adaptation in maize populations. Such approaches can also help identify key genes involved in adaptation to specific ecological niches, thereby informing breeding programs aimed at developing climate-resilient maize varieties.
7.3 Implications for breeding and conservation
The insights gained from gene flow studies have profound implications for maize breeding and conservation. Understanding the genetic basis of adaptation and the impact of gene flow from MVs into LRs and WRs can guide the development of new maize varieties that are both high-yielding and resilient to environmental stresses (Rojas-Barrera et al., 2019; Wisser et al., 2019). Additionally, conservation strategies can be informed by genomic data to maintain and enhance the genetic diversity of maize. This is crucial for preserving the evolutionary potential of maize and ensuring its long-term sustainability as a crop.
7.4 Policy and ethical considerations
The findings from gene flow studies also raise important policy and ethical considerations. The introgression of genes from MVs into traditional LRs and WRs can have both positive and negative effects on genetic diversity and crop resilience (Rojas-Barrera et al., 2019). Policymakers need to balance the benefits of modern breeding techniques with the need to protect indigenous genetic resources. Ethical considerations include the rights of local communities to their traditional varieties and the potential impacts of gene flow on these varieties. Developing policies that promote sustainable agricultural practices and the conservation of genetic diversity will be essential for the future of maize cultivation.
8 Concluding Remarks
This study has provided a comprehensive overview of the evolutionary dynamics of maize, with a particular emphasis on the role of gene flow in shaping its genetic diversity and adaptation. Various aspects of gene flow were explored, from its impact on genetic diversity, agronomic traits, and pest and disease resistance, to its implications for conservation and breeding strategies. The findings underscore the significant influence of gene flow from modern maize varieties (MVs) to traditional landraces (LRs) and wild relatives (WRs), highlighting its contribution to genetic adaptability and resilience in maize.
The studies discussed illustrate how gene flow facilitates the integration of beneficial alleles into maize populations, enhancing their ability to adapt to changing environmental conditions. The incorporation of genes from MVs into LRs and WRs has not only increased genetic diversity but also led to the development of new varieties with improved agronomic and resistance traits. Furthermore, the adaptive introgression events have provided crucial insights into the evolutionary potential of maize, allowing for rapid adaptation to diverse ecological niches.
Looking forward, the role of gene flow in maize evolution appears increasingly pivotal as the world faces global challenges such as climate change and the need for sustainable agricultural practices. Gene flow stands as a key mechanism by which maize can continue to adapt and thrive in diverse environments. It will be crucial in developing varieties that can withstand environmental stresses and meet the demands of growing global populations.
Understanding the complex interactions between gene flow, genetic diversity, and environmental factors will be essential for designing effective breeding programs. These programs must not only focus on enhancing yield and resistance traits but also on conserving the genetic heritage of maize. Policies and ethical considerations will play a significant role in guiding these efforts, ensuring that the benefits of gene flow are realized while minimizing potential negative impacts on indigenous maize varieties and their ecosystems.
In conclusion, gene flow is integral to the ongoing evolution and sustainability of maize as a global staple crop. Embracing the dynamics of gene flow, while carefully managing its ecological and genetic impacts, will be key to future maize breeding and conservation strategies, ensuring that this vital crop continues to serve as a cornerstone of food security worldwide.
Acknowledgments
The authors gratefully acknowledges the insightful feedback provided by two anonymous peer reviewers, which greatly contributed to the improvement of this manuscript.
Conflict of Interest Disclosure
The authors affirm that this research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Calfee E., Gates D., Lóránt A., Perkins M., Coop G., and Ross-Ibarra J., 2021, Selective sorting of ancestral introgression in maize and teosinte along an elevational cline, PLoS Genetics, 17(10): e1009810.
Egan P., Muola A., and Stenberg J., 2018, Capturing genetic variation in crop wild relatives: An evolutionary approach, Evolutionary Applications, 11(8): 1293-1304.
Gates D.J., Runcie D., Janzen G., Navarro A., Willcox M., Sonder K., Snodgrass S., Rodríguez-Zapata F., Sawers R., Rellán-Álvarez R., Buckler E., Hearne S., Hufford M., and Ross-Ibarra J., 2019, Single-gene resolution of locally adaptive genetic variation in Mexican maize, bioRxiv, 18: 706739..
ht tps://doi.org/10.1101/706739
Haberer G., Kamal N., Bauer E., Gundlach H., Fischer I., Seidel M., Spannagl M., Marcon C., Ruban A., Urbany C., Nemri A., Hochholdinger F., Ouzunova M., Houben A., Schön C., and Mayer K., 2020, European maize genomes highlight intraspecies variation in repeat and gene content, Nature Genetics, 52: 950-957.
https://doi.org/10.1038/s41588-020-0671-9
He F., Pasam R., Shi F., Kant S., Keeble-Gagnère G., Kay P., Forrest K., Fritz A., Hucl P., Wiebe K., Knox R., Cuthbert R., Pozniak C., Akhunova A., Morrell P., Davies J., Webb S., Spangenberg G., Hayes B., Daetwyler H., Tibbits J., Hayden M., and Akhunov E., 2019, Exome sequencing highlights the role of wild-relative introgression in shaping the adaptive landscape of the wheat genome, Nature Genetics, 51: 896-904.
https://doi.org/10.1038/s41588-019-0382-2
Hu H., Crow T., Nojoomi S., Schulz A., Hufford M., Flint-Garcia S., Sawers R., Rellán-Álvarez R., Estévez-Palmas J., Ross-Ibarra J., and Runcie D., 2022, Allele-specific expression reveals multiple paths to highland adaptation in maize, Molecular Biology and Evolution, 39(11): msac239.
https://doi.org/10.1093/molbev/msac239
Hufford M., Seetharam A., Woodhouse M., Chougule K., Ou S., Liu J., Ricci W., Guo T., Olson A., Qiu Y., Coletta R., Tittes S., Hudson A., Marand A., Wei S., Lu Z., Wang B., Tello-Ruiz M., Piri R., Wang N., Kim D., Zeng Y., O'Connor C., Li X., Gilbert A., Baggs E., Krasileva K., Portwood J., Cannon E., Andorf C., Manchanda N., Snodgrass S., Hufnagel D., Jiang Q., Pedersen S., Syring M., Kudrna D., Llaca V., Fengler K., Schmitz R., Ross-Ibarra J., Yu J., Gent J., Hirsch C., Ware D., and Dawe R., 2021, De novo assembly, annotation, and comparative analysis of 26 diverse maize genomes, Science, 373: 655-662.
https://doi.org/10.1126/science.abg5289.
Kistler L., Thakar H., VanDerwarker A., Domic A., Bergström A., George R., Harper T., Allaby R., Hirth K., and Kennett D., 2020, Archaeological Central American maize genomes suggest ancient gene flow from South America, Proceedings of the National Academy of Sciences of the United States of America, 117: 33124-33129.
https://doi.org/10.1073/pnas.2015560117
Kong D., Wang B., and Wang H., 2020, UPA2 and ZmRAVL1: Promising targets of genetic improvement of maize plant architecture, Journal of Integrative Plant Biology, 62(4):394-397.
https://doi.org/10.1111/jipb.12873
Lal K., Kumar S., Singh L., Singh H., Singh M., and Kumar A., 2020, Genetic study of certain quantitative traits in maize (Zea mays L.), International Journal of Current Microbiology and Applied Sciences, 9: 2508-2513.
https://doi.org/10.20546/ijcmas.2020.905.287
Ma J., Cao Y., Wang Y., and Ding Y., 2022, Development of the maize 5.5K loci panel for genomic prediction through genotyping by target sequencing, Frontiers in Plant Science, 13: 972791.
https://doi.org/10.3389/fpls.2022.972791
Moreno-Letelier A., Aguirre-Liguori J., Piñero D., Vázquez-Lobo A., and Eguiarte L., 2020, The relevance of gene flow with wild relatives in understanding the domestication process, Royal Society Open Science, 7(4): 191545.
https://doi.org/10.1098/rsos.191545
Rojas-Barrera I., Wegier A., González J., Owens G., Rieseberg L., and Piñero D., 2019, Contemporary evolution of maize landraces and their wild relatives influenced by gene flow with modern maize varieties, Proceedings of the National Academy of Sciences, 116: 21302-21311.
https://doi.org/10.1073/pnas.1817664116
Stitzer M., and Ross-Ibarra J., 2018, Maize domestication and gene interaction, The New Phytologist, 220(2): 395-408.
https://doi.org/10.1111/nph.15350
Wijayasekara D., and Ali A., 2021, Evolutionary study of maize dwarf mosaic virus using nearly complete genome sequences acquired by next-generation sequencing, Scientific Reports, 11(1): 18786.
https://doi.org/10.1038/s41598-021-98299-9
Wisser R., Fang Z., Holland J., Teixeira J., Dougherty J., Weldekidan T., Leon N., Flint-Garcia S., Lauter N., Murray S., Xu W., and Hallauer A., 2019, The Genomic Basis for Short-Term Evolution of Environmental Adaptation in Maize, Genetics, 213(4): 1479-1494.
https://doi.org/10.1534/genetics.119.302780
Xu G., Lyu J., Li Q., Liu H., Wang D., Zhang M., Springer N., Ross-Ibarra J., and Yang J., 2020, Evolutionary and functional genomics of DNA methylation in maize domestication and improvement, Nature Communications, 11(1): 5539.
https://doi.org/10.1038/s41467-020-19333-4




. PDF(670KB)
. FPDF(win)
. FPDF(mac)
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Delong Wang
. Yunchao Huang
Related articles
. Maize
. Gene flow
. Genetic diversity
. Adaptation
. Conservation strategies
Tools
. Email to a friend
. Post a comment